
Men are on average taller and more robust than women, and the same is true for males and females in almost all of our domesticated animal species and in most of the captive species we find in zoos and wildlife parks. Given this, one might be tempted to conclude that being larger than one's mate is a general feature of animal masculinity, but this is far from the truth. I recently cataloged sexual differences across the entire animal kingdom and came to a very different conclusion (1). Among animals other than mammals and birds, it is far more common for females to be larger than males. Females are the larger sex in most reptiles other than lizards and crocodilians; in most amphibians and fishes; in the vast majority insects, spiders and other arthropods; in almost all "wormy" animals; in various phyla of tiny animals such as water bears and rotifers; and in almost all parasitic animals. In hard-shelled animals such as molluscs and lamp shells the sexes seldom differ in size, but when they do, females are the larger sex, and the same is true for brittle stars, sea lilies and sea stars. Overall, I discovered size differences between the sexes in 49 different animal classes and in 86 percent of these females were the larger sex (Figure 1).
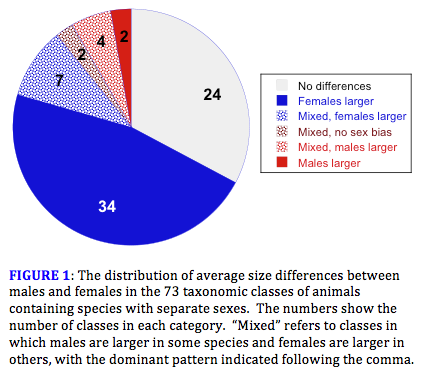
Human males average about 8 percent taller and 20 percent heavier than females, a noticeable but fairly modest difference in size (2). Much greater sexual differences can be found in many other species (3). Among those with males larger than females, the undisputed champion is a small cichlid fish called Lamprologus callipterus in which the males average 12 to 13 times heavier than their mates (Figure 2). Among mammals, southern elephant seals (Mirounga leonina) hold the record, with breeding males weighing as much as seven to eight times more than females. Great bustards (Otis tarda) hold the record for birds, with a weight ratio close to three. These size differences seem huge, but they are trivial when compared to the differences found in species where females are the larger sex. In some of these species, females are more than 10 times longer and weigh 100 to 500,000 times more than males. In such species, adults of the two sexes are often so different that they would never be placed in the same species if they were not observed mating (Table A).

Why do males and females so often differ in size? The answer to this question lies in the different strategies males and females use to ensure their reproductive success. Most female animals produce tens to hundreds-of-thousands of eggs at a time and these eggs grow and mature within the female's body until they are ready to be spawned or laid (4). Within any given species, larger females tend to produce more eggs than smaller females and as a result, they have greater reproductive success (5). This is typical of most reptiles, amphibians, fishes, insects and spiders, and of species in almost all other invertebrate classes, and seems to be the main reason why females are on average larger than males in so many of these classes. The correlation between female size and offspring number tends to be weaker in birds and mammals, which partly explains why females are seldom the larger sex in these classes (6).
TABLE A: Some examples of animal species with extreme sexual disparities in body size.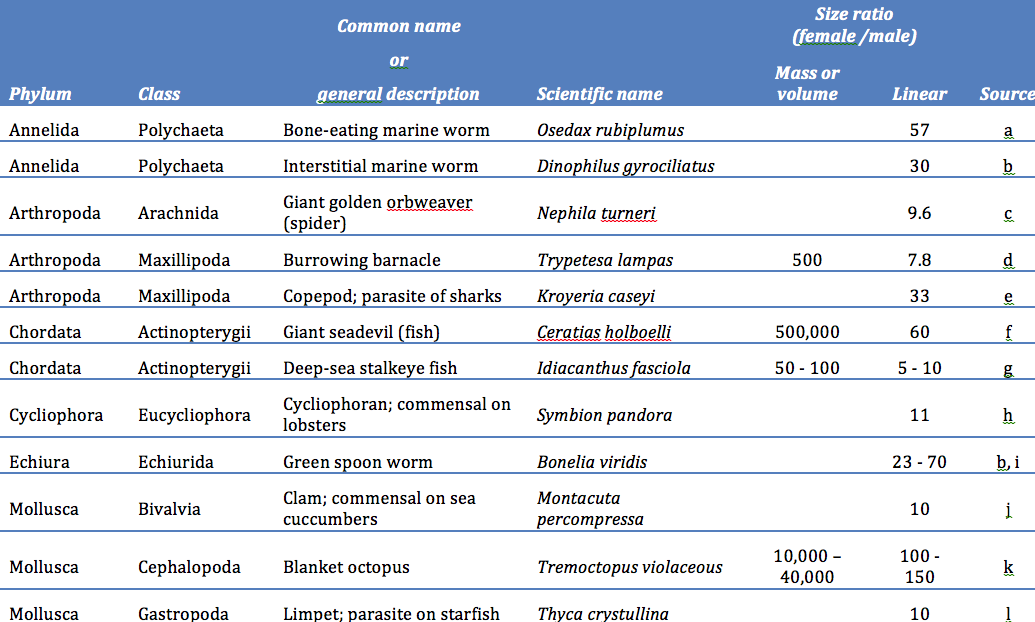
Table sources: a. Rouse et al. (2004), b. Giese & Pearce (1975b), c. Kuntner and Coddington (2009), d. Gotelli and Spivey (1992), e. Benz and Deets (1986), f. Bertelsen (1951), g. Clarke (1983), h. Kristensen (2002) and Obst and Funch (2003), i. Greenwood and Adams (1987), j. Chanley and Chanley (1970), k. Norman, Paul, Finn and Tregenza (2002), l. Elder (1979)
Body size affects male reproductive success as well, but for very different reasons. Males produce offspring only indirectly by fertilizing the eggs of females. In species that occur at relatively high densities or form breeding aggregations, competition among males for mating opportunities is often intense. Males may compete with each other by fighting and physical intimidation or by attempting to attract females with odors (called pheromones) and flamboyant displays. Most often both tactics are used to some extent. In these competitions, larger males often have the advantage over smaller competitors (7). If this size advantage is strong and combined with a weak relationship between female reproductive success and body size, males are likely to be the larger sex. This is the case for the elephant seals, great bustards and cichlid fishes mentioned above, and it is the fundamental explanation for the predominance of male-larger size differences in birds and mammals.
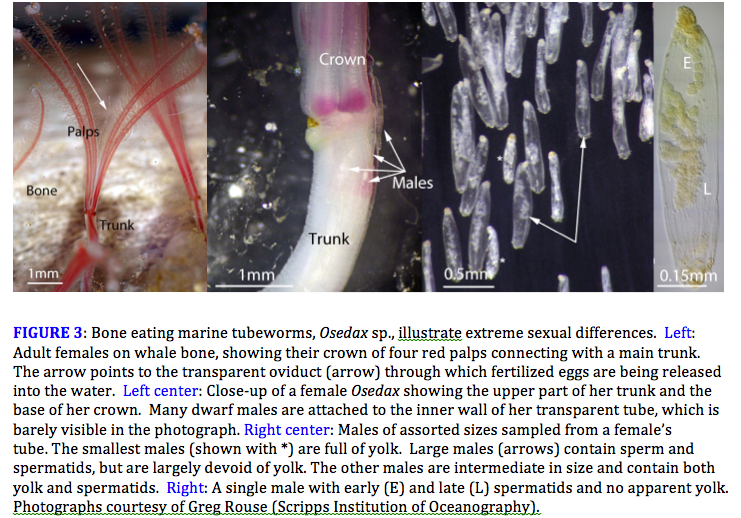
In species that exist at low densities and do not form breeding aggregations, competition among males for mating opportunities is much less intense and may even be absent altogether. The limiting factor for males in these species is the rarity and elusiveness of potential mates rather than the abundance of competitors, and the challenge is simply to find a female. Males is such species must be able to search efficiently for mates and they must be ready to mate whenever they encounter a receptive female. Their signature traits are small size and rapid sexual maturation. The greatest sexual differences occur in species where males adopt this strategy. In the extreme, males are reduced to tiny dwarfs and mature so early that they convert the yolk from their natal egg directly into sperm, bypassing adult morphology entirely (Figure 3).
Although this short summary paints sexual differences with a very broad brush, the overall messages are clear. Females are larger than males in the majority of animal species, and not infrequently they are very much larger. The size difference in humans is modest, at best, when compared to many other animal species, and although we conform to the mammal-bird pattern of males being the larger sex, this is clearly the exception rather than rule. Most animal males are small and quick rather than large and robust, and females are often giants by comparison.
Notes:
1. These sexual differences are summarized in Fairbairn (2013) and full descriptions for each animal class can be found here.
2. This estimate of the ratio of heights is based on the average for heights of males and females in 97 human populations (Holden And Mace 1999), and the World Health Organization growth reference charts for 2007. The weight ratio is for the USA only and is from the Centers for Disease Control.
3. See tables 11.2 and 11.3 in Fairbairn (2013)
4. Females in some species also brood their eggs within their bodies, releasing them only once they have hatched.
5. For more information and examples of the relationship between size and reproductive success in females see Clutton-Brock (1988), Roff (1992), Fairbairn et al (2007) and Fairbairn (2013).
6. Female birds and mammals produce many fewer eggs or offspring at a time and their reproductive success depends strongly on extended parental care. In addition, although female birds lay clutches of eggs, they produce and lay only one egg at a time, so never hold entire clutches of eggs within their bodies. Thus, the volumetric constraints on egg number that may apply to females in other classes are absent in birds.
7. For more information about the influence of body size on male competition for mates, see Andersson (1994), Fairbairn et al (2007), Fairbairn (2013)
References:
Andersson, M. 1994. Sexual Selection. Princeton University Press, Princeton, NJ.
Benz, G. W., and G. B. Deets. 1986. Kroyeria caseyi sp.nov. (Kroyeriidae: Siphonostomatoida), a parasitic copepod infesting gills of night sharks (Carcharhinus signatus (Poey, 1868)) in the western North Atlantic. Can. J. Zool. 64:2492-2498.
Bertelsen, E., ed. 1951. The Ceratioid Anglerfishes. Ontogeny, distribution and biology. Bianco Luno, Copenhagen.
Chanley, P., and M. H. Chanley. 1970. Larval development of the commensal clam, Montacuta percompressa Dall. Proc. malac. Soc. Land. 39:59-67.
Clarke, T. A. 1983. Sex ratios and sexual differences in size among mesopelagic fishes from the Central Pacific Ocean. Marine Biology 73:203-209.
Clutton-Brock, T. H. e. 1988. Reproductive Success. University of Chicago Press, Chicago.
Elder, H. Y. 1979. Studies on the host parasite relationship between the parasitic prosobranch Thyca crystallina and the asteroid starfish Linckia laevigata. J. Zool. Lond. 187:369-391.
Fairbairn, D. J. 2013. Odd Couples. Extraordinary Differeneces between the Sexes in the Animal Kingdom. Princeton University Press, Princeton and Oxford.
Fairbairn, D. J., W. U. Blanckenhorn, and T. Székely, eds. 2007. Sex, Size and Gender Roles. Evolutionary Studies of Sexual Size Dimorphism. Oxford University Press, Oxford, UK.
Giese, A. C., and J. S. Pearse, eds. 1975. Reproduction of marine Invertebrates, Volume III Annelids and Echiurans. Academic Press, NY.
Gotelli, N. J., and H. R. Spivey. 1992. Male parasitism and intrasexual competition in a burrowing barnacle. Oecologia 91:474-480.
Greenwood, P. J., and J. Adams. 1987. The Ecology of Sex. Edward Arnold, London, UK.
Holden, C., and R. Mace. 1999. Sexual Dimorphism in Stature and Women's Work: A Phylogenetic Cross-Cultural Analysis. American Journal of Physical Anthropology 110:27-45.
Kristensen, R. M. 2002. An introduction to loricifera, cycliophora, and micrognathozoa. Integrative and Comparative Biology 42:641-651.
Kuntner, M., and J. A. Coddington. 2009. Discovery of the Largest Orbweaving Spider Species: The Evolution of Gigantism in Nephila. PloS One 4.
Norman, M. D., D. Paul, J. Finn, and T. Tregenza. 2002. First encounter with a live male blanket octopus: the world's most sexually size-dimorphic large animal. New Zealand J. Marine and Freshwater Res. 36:733-736.
Obst, M., and P. Funch. 2003. Dwarf male of Symbion pandora (Cycliophora). J. Morphol. 255:261-278.
Roff, D. A. 1992. The Evolution of Life Histories. Chapman and Hall, New York, NY.
Rouse, G. W., S. K. Goffredi, and R. C. Vrijenhoek. 2004. Osedax: Bone-eating marine worms with dwarf males. Science 305:668-671.