Why do we believe fictions in the face of facts? When do the scales fall from our eyes?
These are the questions that dog me in relation to modern US political discourse, but also A. J. Bateman's hyper-cited 1948 experimental study in fruit flies of reproductive competition (a.k.a. sexual selection). Sexual selection is the Darwinian process that results in the evolution of peacock's tails and other fancy male traits, and that may account for the evolution of other sex differences.
At the turn of the century, I read Bateman's influential study with a different eye than when I read it as a graduate student in 1972. In that interval the study continued to provide an adaptive justification for good girl feminine neediness (good girl flies) and bad boy ardent promiscuity (randy and rapacious boy flies) as evolutionarily adaptive, sex-specific behavioral alternatives resulting from sexual selection. The evolutionary justification was that males' profligacy earned them fitness rewards, but that females gained no adaptive benefits from mating with more than one male.
I wondered if an alternative explanation wouldn't also explain Bateman's data: Did ecological and social opportunities and constraints that individuals inevitably experience explain the sex differences Bateman saw? And, then I wondered why his study had never been replicated.
Bateman had figured out who mated with whom and how many offspring each of his subjects had by looking at the dramatic visible mutations that offspring inherited from their parents. The modern tests we proposed to use are like the genetic tests that can prove whether you are the father of your wife's son Jack or her daughter Jill.
My eye scales were rubbed off after my collaborator Wyatt Anderson, convinced me that we should not only use modern tools but also really repeat Bateman using his exact, old-fashioned, labor-intensive method of odd-looking subjects. And besides, we realized that no one else had yet published a repetition of this classic experiment.
The rub came as I was reading Bateman's paper with an eye to repetition when I focused on his table 4 (below) showing the only complete set of observations as he first observed them for a single population out of his 64 experimental populations. Three adult females and three adult males were the subjects whose mate number and number of offspring (reproductive success, RS) Bateman wished to count.
The devil is in the details, so I have to ask you too to look at the table below to see what Bateman saw. The top row of the table contains the subject females by their "nametags", their visibly obvious phenotypic oddities that were coded by genes unique-to-each-adult. The far right column contains the subject males by their nametags.
The numbers in each cell are counts of the number of offspring for each possible combination of parents.
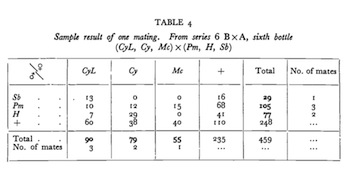
Reprinted by permission from Macmillan Publishers Ltd. License Number: 2960430598991 from Bateman, A. J. 1948. Heredity (Edinb) 2:349-368, p 357.
The first column shows that the female with the CyL nametag had 13 + 10 + 7 = 30 offspring each of whom carried CyL's curly wings and notched eyes, 13 of whom also inherited their father's dramatically shortened body bristles (nametag: Sb), 10 others had their dad's brown eyes and darkened body (nametag: Pm), and 7 were hairless over much of their bodies and had bald heads like their dad's (nametag: H). The CyL female also had 60 kids whose paternity was unobservable because each of them inherited their dad's wild-type gene, indicated with a + sign instead of his nametag gene meaning that for CyL+ kids there was no way to know who was the daddy. Similarly the rows show for each male how many females each mated and how many offspring each had with each of his mates. To count up the number of mates for each female subject (totals are in the final column) and each male subject (totals in the bottom row), you should add up each time their nametag appears in the offspring along with a nametag of one of the opposite sex subjects. In this population one female had 3 mates, another 2, and another only 1. Ditto for the males.
Simple and brilliant, huh?
Yes, except that the data in Bateman's table 4 rock the foundations of Bateman's claims. The attentive high school biology student - with just the data in the table above and a modest understanding of Mendel's laws - can question the reliability of Bateman's method and the veracity of his conclusions.
Here's what you need to know: each adult subject carried only one visible mutation - it's unique nametag - on only one of the two chromosomes of its nametag locus, but a nondescript gene (a "wildtype" gene that codes for the usual-looking fly) on the other and also had only wildtype genes at each of the chromosomes of everyone else's nametag loci. We call such individuals "heterozygote dominants". This meant that each adult looked to our eyes distinctively different. Each had a nametag phenotype. The underlying genetics that produced distinct adult phenotypes also means that each adult produced only two kinds of gametes, i.e., sperm (if he's a male) or ova (if she's a female): One type with the parental nametag (the marker gene, like an Sb or a Cy) and one type without the nametag (a wild-type gene, a +).
Remember that gametes carry one chromosome and that gametes join forces during fertilization to produce a new embryo with two chromosomes.
Now you can calculate what Mendel's rules say about the expected frequency of parental nametags in a set of offspring.
Set up a 2x2 table like the one in the Mexican bark painting below - a cartoon of the simple math problem of reproduction, reflected in Mendel's rules. In the upper left you can see a male fly, a potential father with notched wings (his nametag) and a female fly, a potential mother with stripped eyes (her nametag). If these two adults mated and if there were no effects on the survival of the offspring from inheriting notched wings and stripped eyes together, these parents would produce four types of offspring in equal frequencies: 25% would inherit their mom's stripped eyes and their pop's notched wings (upper left hand cell of the matrix), ¼ their mom's stripped eye but her nondescript (i.e., "wildtype") wings (bottom left cell of the matrix), ¼ with nondescript eyes, but their pop's notched wings (upper right hand cell), and ¼ with neither of their parent's phenotypic markers. And, this means that for every pair of parents in the population 1/4 of their offspring would have two nametags, 1/4 only their dad's nametag, 1/4 only their mom's nametag, and 1/4 neither nametag, giving us an easy way to test whether Bateman's results conform to Mendel's rules.

Mendel's expectations of equal frequency of offspring phenotypes given that parents are heterozygote dominants each at a different and unique genetic locus. Image of acrylic and black ink on amate paper with permission of artist Sergio Castrezana©.
Now look again at Bateman's table. Doing a simple statistical test for the equality of frequencies of the four expected types of offspring (those with two nametags, only a nametag from a dad, only a nametag from mom, or no nametags at all) shows that the data in Bateman's table violate the basic assumption on which the fairness of Bateman's data and the reliability of his conclusions depended.
Oops!
Part of the sample informing how many mates each adult had is missing. Invisible matings meant that any counts of adults with zero mates were inflated and counts of those with 1, 2 and 3 mates under-estimated.
Would those missing 29 offspring have told us that additional females besides lucky CyL also mated with all the males in the bottle? And, were the other 63 populations in Bateman's study similarly biased?
A strict repetition of Bateman's study seemed the only way forward.`